5 Les gaz et l'exercice
5.1 Evolution de la consommation d'oxygène
Nous savons que l'organisme prélève de l'oxygène dans l'atmosphère et rejette du dioxyde de carbone. Ces échanges de gaz sont respectivement appelés : consommation d'oxygène (VO2) et rejet de CO2 (VCO2). Des techniques de récupération des gaz permettent de mesurer les échanges au cours de l'exercice. Les unités utilisées pour les quantifier expriment un volume en fonction d'un temps ou d'une distance. Les plus usités sont le litre par minute (l/min), le millilitre par minute et par kilogramme de poids corporel (ml/min/kg) et le litre par kilomètre (l/km).
 VO2 et vitesse de course
VO2 et vitesse de course
Au repos, nous utilisons une certaine quantité d'oxygène. Quand nous faisons un effort, cette quantité s'accroît. Lors de l'effort progressif, la consommation d'oxygène (VO2) augmente de manière linéaire avec l'intensité de l'exercice (Astrand et Rodahl, 1994) jusqu'à un niveau caractérisé par une absence d'augmentation de l'oxygène consommé malgré une majoration de la charge de travail. Ce niveau correspond à la consommation maximale d'oxygène (VO2max).
| |
 |
|
| |
Représentation de l'évolution de la VO2 en fonction de la vitesse de course
|
|
Au-delà de VO2max toute énergie supplémentaire provient exclusivement de la filière acide.
La représentation ci-dessus est largement idéalisée. Les mesures de VO2 sur le terrain, pendant un test progressif de course (paliers de 3 minutes avec 1 minute d'arrêt entre chaque vitesse) donne une évolution beaucoup plus saccadée.
| |
 |
|
| |
Evolution de la VO2 d'un coureur de demi-fond en fonction de la vitesse de course au cours d'un test progressif par paliers de 3' avec 1' d'arrêt entre chaque palier
|
|
VO2 et durée de l'effort
| |
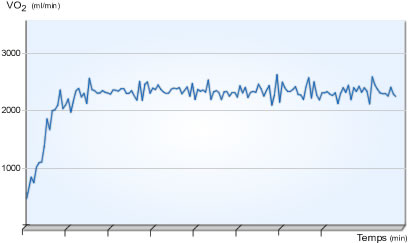 |
|
| |
Représentation de l'évolution moyenne de la VO2 en fonction du temps au cours d'un exercice réalisé à vitesse constante
|
|
La VO2 augmente très rapidement dès le début de l'exercice (composante rapide) puis plus lentement (composante lente) jusqu'à atteindre un état d'équilibre ou alors son niveau maximal (Di Prampero et al 1970, Whipp et Wasserman 1972).
Pour des intensités inférieures à la vitesse maximale aérobie (VMA), le temps mis pour atteindre l'état d'équilibre est d'autant plus grand que l'intensité est importante (Whipp et al 1982...). A l'approche et au-dessus des intensités correspondantes à VO2max, plus la puissance de l'exercice est élevée, plus l'augmentation de la VO2 est rapide (Astrand et Saltin 1961 in Astrand et Rodahl, 1994).
Tests de course
Plusieurs protocoles permettent de mesurer la consommation d'oxygène associée aux différentes vitesses de course. Généralement, ces épreuves sont effectuées en laboratoire. L'athlète court alors sur un tapis roulant. Toutefois une nouvelle génération d'analyseurs de gaz portables permet de courir sur la piste ou sur un parcours balisé.
Dans tous les cas, les grands traits de ces tests correspondent à ceux que nous avons donnés dans le secteur tout sur l'entraînement .
Les mesures effectuées renseignent le coureur sur trois notions fondamentales pour son entraînement :
 VO2max
VO2max
La validité de VO2max est basée sur la forte corrélation la liant au niveau de performance du 800m au marathon (Costill et al 1973, Foster 1983). Au sein d'une grande population, VO2max est un critère discriminant de l'aptitude aérobie. Plus un athlète a une consommation d'oxygène importante, plus il a de chances de réaliser de bon temps en course.
Pourtant, VO2max n'explique pas tout. Le coureur Dereck Clayton a réalisé 2h08' au marathon avec une VO2max de 69,7ml/kg/min. Selon "la théorie extrapolée de VO2max", il n'aurait jamais dû réaliser mieux que 2h23'. A l'inverse, cette même théorie attribuait un temps inférieur à 2h04' à l'infortuné Gary Tuttle (VO2max 82,7ml/kg/min) qui a dû se contenter d'un modeste 2h17' !
Le problème est que l'on a extrapolé au plus haut niveau des résultats trouvés sur des groupes de niveaux différents. Lorsque l'analyse porte sur un niveau élevé homogène plus rien ne tient. Dans une étude portant sur un groupe de coureurs de niveau très homogène, Conley et Krahenbuhl (1980) ne trouvent pas de rapports entre VO2max et la performance sur 10000 mètres. Par contre, dans ce même groupe, l'efficacité de la foulée était un excellent indice de la performance en course à pied. Une VO2max élevée est donc une condition nécessaire mais insuffisante pour prétendre à un haut niveau de performance d'où l'intérêt de disposer d'un indicateur ajoutant l'aspect technique au critère physiologique.
vVO2max ou VMA
La vitesse à la consommation maximale d'oxygène (vVO2max) nommée aussi vitesse maximale aérobie (VMA) est la première vitesse à partir de laquelle VO2 n'augmente plus significativement. Elle réalise la réunion de deux paramètres essentiels de la performance en course de durée : la VO2max et l'efficacité de la foulée. Selon Mercier et Léger (1986) cette double détermination lui conférerait un meilleur degré de prévision de la performance que tous les autres prédicateurs pris isolément.
Pour l'entraînement, la VMA est l'étalon à partir duquel les vitesses de course sont déclinées. On parlera, par exemple, d'une séance de 6x2' à 95% de VMA. Dans le secteur "tout sur l'entraînement" (tout sur l'entraînement) nous avons séparé le secteur aérobie en 3 zones d'allures. La zone verte couvre les vitesses comprises entre 50% et 75% de VMA, la jaune va de 75 à 90% de VMA alors que l'orange s'étend de 90 à 105% de VMA.
Bien que VMA soit l'expression du coût énergétique de la course à cette vitesse, la détermination de ce coût n'est pas évident compte tenu du fait que l'exercice est réalisé pour partie en acidose. C'est pourquoi il est utile d'enregistrer la VO2 au cours d'un exercice continu réalisé à un niveau aérobie stabilisé.
vitesse à VO2 sous maximale : l'économie de course
En 1982, Cavanagh et Williams ont défini l'économie de course comme le niveau de VO2 à une vitesse sous maximale donnée. La comparaison dans le temps des VO2 d'un coureur à une même vitesse sous maximale permet de savoir si son efficacité en course s'est améliorée.
Pour calculer l'économie de course, avant le test de VO2max, les coureurs s'échauffent pendant 5', récupèrent 5' et courent alors 5 à 10 minutes à 14km/h. La VO2 moyenne est calculée dans la deuxième partie de la course, ce qui permet d'assurer sa stabilité (Astrand et Rodahl, 1994). Elle est prise comme référence de l'économie de course de l'athlète.
Enfin, signalons que la prise en compte de certains indicateurs physiologiques permettrait de repérer l'allure ou la zone d'allures associée à un passage accrue à l'utilisation de l'énergie d'origine anaérobie.
Tous ces indices nous donnent accès au niveau de développement et d'efficacité de la filière aérobie du sujet. Le suivi des valeurs dans le temps apporte une information sur l'incidence des exercices d'entraînement sur cette filière et sur l'économie de course des athlètes. VO2 a donc fonction de contrôle et de prédiction des effets de l'entraînement.
Effets d'entraînement
sur VO2max
Les effets de l'entraînement sur la consommation d'oxygène peuvent être évalués en comparant des groupes de sujets plus ou moins entraînés (sédentaires / haut niveau) ou en suivant des athlètes au cours d'un processus d'entraînement. Dans les deux cas, l'entraînement en aérobie conduit à une augmentation de la VO2max. Les athlètes qui atteignent le plus haut niveau mondial présentent des valeurs deux fois plus élevées que celles d'un sujet moyen. Les valeurs maximales relevées chez les hommes sont supérieures à 90 ml/min/kg chez les hommes et 75 chez les femmes. La moyenne de la population se situe autour de 40ml/min/kg (à propos du rapport entre le niveau atteint et VO2max, voir le secteur tout sur l'entraînement).
sur la transition
Pour un niveau donné d'exercice, l'entraînement améliore la vitesse d'échange des gaz entre l'organisme et l'environnement (Norris et Petersen, 1998). Ce progrès se traduit par une élévation plus rapide de la consommation d'oxygène, suite à une demande énergétique brusquement accrue, chez les athlètes possédant la meilleure aptitude aérobie (Powers et al, 1985).
sur l'économie de course
A un niveau d'effort donné, la VO2 est moins élevée après entraînement qu'avant (Billat et al, 1999). Ce constat semble être le reflet d'une amélioration de l'économie de course (à ce propos, voir le secteur biomécanique).
Nous verrons dans le paragraphe ce qui, au niveau physiologique, permet d'expliquer de telles améliorations. Mais avant, disons un mot sur d'autres paramètres respiratoires.
5.2 Autres échanges gazeux
Le volume de dioxyde de carbone rejeté par l'organisme Au cours de l'exercice progressif VCO2 augmente tout d'abord de façon linéaire avec l'augmentation de l'intensité. A partir d'un certain niveau d'effort, la courbe d'évolution de VCO2 marque un point de rupture signifiant un rejet de CO2 plus marqué (figure ci-dessous). Cette évolution est directement liée à l'augmentation du volume d'air expiré (VE) dans la mesure où la fraction de CO2 dans l'air expiré n'augmente jamais au-dessus de 4% (Monod et Flandrois, 1994).
Le volume d'air
L'exercice d'intensité progressivement augmentée s'accompagne d'un accroissement linéaire du volume d'air expiré (débit ventilatoire = VE) jusqu'à un niveau variable selon les individus. Au-delà de ce point, le VE s'élève de manière plus prononcée, marque d'une hyperventilation relative d'exercice (figure suivante). Une rupture dans l'évolution du débit ventilatoire se produirait deux fois au cours du passage de l'exercice modéré à l'exercice maximal aérobie (Wasserman 1984 ; Skinner et Mc Lellan 1980).
| |
 |
|
| |
Représentation de l'évolution de la consommation d'oxygène (VO2), de la ventilation (VE) et du dioxyde de carbone rejeté (VCO2) en fonction de la vitesse de course
|
|
Le point de décrochage pour lequel VE augmente plus rapidement que la VO2 a été identifié par Hollman (1959) comme un "point d'efficience ventilatoire optimale". En 1964, Wasserman et al ont donné le nom de seuil anaérobie (anaerobic threshold) à l'intensité de l'effort repérée par le décrochage de l'évolution du rapport VE/VO2. Ce seuil est actuellement utilisé par de nombreux chercheurs sous le nom de seuil ventilatoire. Il a été souvent rapproché du seuil lactique
Au niveau de la pratique ces données semblent indiquer que :
l'allure seuil, c'est à dire la vitesse associée au footing rapide, assure une ventilation ample et efficace. Elle correspond à un niveau d'échange gazeux optimal.
au-delà de cette vitesse la ventilation deviendrait de plus en plus saccadée et de moins en moins profonde. Cette tendance à ventiler rapidement et de manière superficielle pourrait nuire à la performance.
Pour pouvoir s'assurer de cette dernière hypothèse, nous devons passer d'une analyse qui se cantonnait aux gaz mesurés au niveau de la bouche à une approche prenant en compte les gaz présents dans le sang. Ce faisant, nous seront en mesure d'accéder à certaines limites de l'exercice physique.
5.3 Oxygène et limites de l'exercice
Présentation
Nous tirons la grande majorité de notre énergie de processus de transformation ayant l'oxygène comme accepteur final. Sans cet élément, nous ne disposerions que de 5% de notre capacité énergétique.
Si l'oxygène est important pour la survie, il l'est également pour la réalisation d'une performance physique. Parmi toutes les causes d'arrêt de l'exercice, l'hypothèse d'un déficit d'oxygène est largement répandue.
Cette conjecture fait l'objet d'un grand débat entre spécialistes de la physiologie de l'effort. Deux interrogations sont débattues.
1La capacité à fournir un effort important est-elle limitée par l'apport d'oxygène ?
2Si tel est le cas, où se situe la limitation ?
Nous proposons de prendre le cheval par la queue (le pauvre.) en nous efforçant, dans un premier temps, de répondre à la deuxième question.
Quelles limites à la consommation d'oxygène ?
Pour tirer partie de l'oxygène, l'organisme doit le capter dans l'air, l'apporter jusqu'aux muscles et l'utiliser au sein des mitochondries. Cette utilisation peut être limitée au niveau :
du couloir aérien qui mène de l'atmosphère jusqu'aux alvéoles pulmonaires
du passage entre l'air pulmonaire et le sang (barrière pulmonaire)
du transport par le sang, des poumons aux petites artères situées à proximité des muscles
du passage des capillaires sanguins aux mitochondries contenues dans les cellules musculaires
de l'utilisation de l'oxygène à l'intérieur des mitochondries.
Selon le niveau du vivant auquel s'appliquent ces 5 limites possibles, deux conjectures ont été formulées pour rendre compte d'une limitation de l'apport d'oxygène pendant l'exercice : l'hypothèse centrale et l'hypothèse périphérique.
Dans la première configuration, la capacité de l'organisme à utiliser l'oxygène serait limitée par les trois premières propositions. La ventilation et/ou le passage de la barrière poumon-sang et/ou le transport de l'oxygène dans le sang seraient à l'origine d'une incapacité de l'organisme à amener suffisamment d'oxygène à proximité des muscles. L'objectivation de cette hypothèse passe par la mesure ou l'évaluation du débit cardiaque et du taux d'oxygène dissous dans le sang artériel.
Dans le second cas de figure, la consommation d'oxygène serait limitée par la capacité de cet élément à passer du sang aux mitochondries contenues dans les muscles et/ou à être utilisé au sein de cet organite. En pratique, l'utilisation de l'oxygène par les muscles est évaluée par la mesure de la différence entre les taux d'oxygène mesurés dans les artères (sang arrivant aux muscles) et les veines (sang repartant aux poumons). Les scientifiques parlent de différence artéro-veineuse en oxygène. Sa part peut également être évaluée à partir de la connaissance du débit sanguin et de la VO2 mesurée au niveau de la bouche.
Pour nous faire une idée sur ces deux hypothèses, voyons ce qui se passe aux endroits incriminés lorsque la consommation d'oxygène plafonne c'est à dire lorsque l'exercice est effectué à une intensité supérieure ou égale au maximum aérobie.
5.3.1 Facteurs centraux
La ventilation
Nous avons vu que la ventilation augmentait d'abord proportionnellement puis plus rapidement que l'intensité d'exercice. Le volume d'air déplacé à chaque cycle respiratoire et la fréquence de ces cycles participent à cette élévation.
Quand le niveau d'exercice atteint puis dépasse la consommation d'oxygène, la ventilation continue à augmenter. Elle peut ainsi atteindre des valeurs proches de 200 litres d'air mobilisés à chaque minute. La quantité d'oxygène prélevée à chaque inspiration ne chute pas par rapport aux valeurs de repos.
Ces paramètres semblent indiquer que la ventilation ne limite pas la prise d'oxygène par l'organisme. Des calculs théoriques prévoient même que si tous les systèmes d'utilisation d'oxygène étaient au niveau de la ventilation, des VO2max trois fois supérieures à celles observées devraient être observées.
La ventilation paraissant innocentée voyons le cas du passage du poumon au sang
La barrière entre le poumon et le sang
A l'exercice, la surface de contact entre les alvéoles pulmonaires et le sang ainsi que la perméabilité de la surface des poumons sont accrues. La vitesse de transfert des gaz à travers la paroi pulmonaire est accélérée.
Même si sur ce point les polémiques sont vives, il semblerait que l'accroissement de cette vitesse soit proportionnel à l'effort fourni et ne présente pas de limites pour des intensités élevées.
Si tel est le cas, la capacité de l'oxygène à passer la barrière entre le poumon et le sang ne serait pas en cause dans la limitation de la consommation maximale d'oxygène (VO2max).
Les données sur ce point étant contradictoires, observons ce qui se passe au niveau du sang en partance des poumons.
L'oxygène dans le sang
Nous avons vu que l'hémoglobine capte l'oxygène entrant dans le sang puis le transporte jusqu'aux cellules. Si à la sortie des poumons, la totalité de l'hémoglobine a fixé de l'oxygène, nous pouvons estimer que la barrière pulmonaire ne limite pas le passage de l'oxygène.
Au repos, 97-99% de l'hémoglobine porte de l'oxygène. Nous disons qu'elle est saturée en oxygène à 97-99%. A l'exercice intense, les sujets sédentaires et moyennement entraînés ne montrent pas de baisses du taux de saturation de l'hémoglobine. En revanche, chez certains athlètes très entraînés ce taux baisse jusqu'à atteindre des valeurs proches de 90% (Powers et al 1988). Près de 10% de l'hémoglobine n'a pas le temps de se charger en oxygène pendant le passage au niveau des poumons (Dempsey et al 1984, Williams et al 1986, Powers et al 1988, Nichols et al 1999). La diminution de la capacité de transport de l'O2 qui résulte de cette baisse de la saturation pourrait être un facteur limitant la performance (Ekblom et al 1975, Hughson et al 1995).
Cette éventuelle limitation centrale est confirmée par le fait que tous les moyens permettant d'améliorer l'apport d'oxygène aux muscles (dopage sanguin, ingestion d'EPO, inhalation d'air enrichi en oxygène) contribuent, chez les athlètes confirmés présentant une désaturation sanguine en oxygène, à :
augmenter la VO2max
ramener la saturation en oxygène à ses valeurs maximales (Powers et al, 1989).
En revanche, ces artifices ne changent rien aux valeurs maximales aérobies atteintes par des sujets moins entraînés ne présentant pas de baisse du taux d'oxygène dans le sang.
Pour le sportif moyennement entraîné les échanges gazeux ne constituent donc pas un facteur limitant VO2max. En revanche, chez certains athlètes très entraînés, dont la VO2max est supérieure à 60ml/min/kg, la diffusion entre le poumon et le sang peut limiter la consommation d'oxygène.
Les chercheurs qui ont poussé les investigations ont donné plusieurs explications à ce phénomène. Nous rapportons celle sur laquelle l'athlète est susceptible de pouvoir agir.
Il est possible que la propension à capter l'oxygène puisse être influencée par la respiration adoptée. A l'effort maximal, la fréquence respiratoire augmente au détriment de l'amplitude. L'athlète adopte une respiration plus rapide mais moins ample. Les conséquences d'une telle ventilation seraient doubles :
les poumons s'empliraient moins complètement
les molécules de gaz auraient une vitesse accrue.
Ces ajustements pourraient nuire à la diffusion de l'oxygène dans le sang. Les athlètes entraînés présentant une désaturation du sang en oxygène - et qui le savent - auraient donc tout intérêt à favoriser l'amplitude de leur respiration. Du moins, ne risquent-ils rien à essayer.
Nous tenons donc un candidat susceptible de limiter l'exercice par insuffisance d'oxygène. Voyons si nous trouvons d'autres prétendants.
Le transport de l'oxygène par le sang
L'exercice demande davantage d'oxygène donc de sang aux muscles. Le volume de sang et la vitesse à laquelle il est acheminé à travers l'organisme sont donc susceptibles d'influer sur la capacité à amener l'oxygène aux cellules musculaires.
Nous avons vu qu'au cours de l'effort, le débit cardiaque c'est à dire le volume de sang transporté par l'organisme à chaque minute augmentait d'un facteur 8. Cet accroissement est le résultat de l'élévation de la fréquence cardiaque et d'une large augmentation (x2) de la quantité de sang expulsée du coeur à chaque battement. Une limitation de ce débit - par exemple, suite à l'ingestion d'un bétabloquant - entraîne une baisse de VO2max chez les personnes moyennement entraînées.
Par ailleurs, une baisse du volume sanguin diminue également VO2max dans des proportions égales à la quantité de sang enlevée.
Enfin, les techniques qui contribuent à accroître le volume et le débit sanguin (comme l'auto-transfusion) conduisent à une élévation de la consommation d'oxygène.
Ces remarques laissent à penser que le débit sanguin est susceptible de limiter le transport d'oxygène au moins chez les personnes entraînées.
Il est toutefois difficile de savoir dans quelles mesures le débit et le volume sanguin peuvent constituer une limite aux aptitudes aérobies d'un sportif. Plus exactement, disons que l'évolution de ces deux paramètres ne peut être pensée sans envisager la redistribution du flux sanguin qui accompagne l'effort physique.
Notre organisme se débrouille pour envoyer le sang là où il est indispensable. Pendant l'exercice, les muscles ont besoin des métabolites et des gaz transportés par le sang. Le volume sanguin n'étant pas illimité, pour satisfaire à cette demande, du sang doit être retiré à certains endroits pour être redistribué à d'autres. Pendant l'exercice, le flux sanguin est réduit dans les intestins, le foie, les reins et augmenté à proximité des muscles (de 1l/min au repos à 18l/min à l'effort). Ceci est rendu possible notamment grâce à l'ouverture massive de minuscules vaisseaux sanguin appelés capillaires.
En entrant dans les capillaires, nous arrivons dans le domaine des limitations périphériques.
5.3.2 Facteurs périphériques
Dans le muscle au repos, un capillaire sur 30-40 laisse passer le sang. A l'exercice maximum, tous sont ouverts. Cette adaptation est en partie due à l'élévation du taux de CO2 à proximité des muscles. Le CO2 participe également à une meilleure libération de l'oxygène fixé sur l'hémoglobine (effet Bohr). Grâce au premier effet du CO2, le débit sanguin au niveau des muscles peut monter de 4 à 200 ml de sang/100g de muscle/min.
Cet accroissement du débit sanguin dans les tissus actifs est avec la meilleure libération de l'oxygène un facteur essentiel de l'apport de cet élément aux cellules.
Distance du sang aux cellules
Pour illustrer l'importance qu'occupe la distance entre la source d'oxygène et son lieu d'utilisation, parlons de l'oxyde de carbone. Ce gaz pénètre facilement dans le sang où il prend la place de l'oxygène sur l'hémoglobine. Une inhalation importante de ce gaz provoque la mort rapide des animaux aussi gros que l'homme. Il tue en empêchant l'hémoglobine du sang d'amener de l'oxygène à nos tissus. Or un animal plus petit comme la souris peut survivre à la saturation intégrale de son sang par l'oxyde de carbone. Quelle différence anatomique est susceptible d'expliquer ce phénomène ?
Si la souris survit, c'est parce que suffisamment d'oxygène peut passer à travers sa peau et ses poumons et diffuser librement jusqu'à ses tissus. Si l'homme ne peut réaliser cette prouesse c'est, avant tout que le chemin à accomplir par les molécules d'oxygène est beaucoup trop long pour un processus de transport aussi inefficace que la diffusion.
La distance séparant l'élément qui en dernière instance dispose d'oxygène et l'élément qui en a besoin est une donnée centrale du transport de l'oxygène. Elle pourrait constituer une limite à l'apport d'oxygène aux cellules. Pour Roca et al (1989) VO2max peut être limitée par la capacité de diffusion de l'oxygène des globules rouges présents dans les capillaires jusqu'aux mitochondries musculaires. Un effet immédiat de l'entraînement aérobie est d'ailleurs d'augmenter la micro-circulation à proximité des muscles.
Pourtant, si théoriquement une telle limitation semble s'imposer, elle est difficile à établir par des données expérimentales. Les connaissances que nous avons de la dynamique sanguine à proximité des muscles sont, en effet, extrêmement lacunaires.
Compte tenu de ce que nous venons de dire de la respiration, de la circulation et du transport jusqu'aux cellules, il semble que l'apport d'oxygène au muscle puisse être limité. Est-ce à dire que le muscle peut se trouver en panne d'oxygène ?
5.3.3 Pas assez d'oxygène ?
Des études portant sur les muscles d'animaux montrent que leur pression en oxygène reste toujours suffisamment importante pour assurer les réactions aérobies. Même lors de contractions maximales, le muscle ne se trouverait jamais en manque d'oxygène (Connett et al, 1984).
Tout ce que nous venons de dire sur le transport de l'oxygène n'aurait donc aucune importance ? L'apport d'oxygène ne saurait donc être un facteur limitant son utilisation par le muscle ?
Si tel est le cas, comment interpréter la baisse d'oxygène sanguin observée chez l'athlète ? Comment expliquer les différences de niveau aérobie entre des sédentaires et des athlètes confirmés ? Comment rendre compte de l'augmentation de VO2max accompagnant, par exemple, le dopage sanguin ? Et comment traduire la montée d'acidité accompagnant un exercice intense ?
Quel modèle pourrait rendre compte à la fois d'une présence d'oxygène toujours suffisante au niveau des muscles et d'une limitation de l'organisme à utiliser cet oxygène donc à réaliser un exercice intense sans fatigue due à l'acidité ?
Le modèle abordé dans la page consacrée à l'énergie (physiologie/énergie) conviendrait. Un modèle qui associe les différences de vitesses et la loi d'action de masse.
5.3.4 Un modèle pour l'oxygène
Les différences de vitesses
Nous avons signalé dans les pages consacrées à l'énergie et aux lactates (physiologie/énergie ; physiologie/lactate) que les transformations aérobies se font beaucoup plus lentement que les réactions anaérobies.
Cette capacité à utiliser rapidement l'oxygène présent dans la cellule permet d'expliquer une limite inhérente à l'utilisation de l'oxygène. Cette limite est interne à la mitochondrie.
Des études récentes suggèrent que dans des conditions de flux sanguin maximal, la VO2 de la mitochondrie est limitée par la vitesse du cycle de Krebs (Blomstrand et al 1997). Dès lors, des facteurs comme le nombre et le volume des mitochondries (Dudley et al 1987, Gollnick et al 1985), l'activité maximale des enzymes du cycle de Krebs (comme la Succinate-déshydrogénase - SDH et la citrate synthase (CS) l'oxoglutarate deshydrogénase mais aussi la pyruvate deshydrogénase (Putman et al 1995)) pourraient limiter la capacité du muscle à utiliser l'oxygène.
La principale limitation de VO2max se situerait donc au niveau des mitochondries qui n'arriveraient pas à consommer suffisamment vite l'oxygène pourtant disponible. En parallèle, les processus anaérobies beaucoup plus rapides déverseraient de l'acidité dans la cellule.
Mais si les différences de vitesses étaient la seule cause de fatigue par empoisonnement acide, comment expliquer la désaturation du sang en oxygène observée chez les athlètes confirmés ? Comment expliquer la baisse de performance associée à une diminution du débit sanguin chez les athlètes de niveau intermédiaire ?
L'effet de masse
Cet effet s'applique aussi bien à la transformation d'une substance en une autre qu'au passage d'un endroit à un autre. Prenons l'exemple du transfert d'une molécule. Soit la diffusion d'un produit depuis une zone A vers une zone B. La vitesse de ce transfert est d'autant plus importante que la différence de concentration entre A et B est grande et/ou que la concentration de la substance dans le compartiment A est importante.
Résultats : plus il y a d'oxygène au niveau du poumon, plus il y a de chances que le sang soit saturé. Plus il y a d'oxygène dans le sang, plus son passage dans le muscle sera aisé. Plus la quantité d'oxygène musculaire est grande, plus il est pensable que les réactions aérobies soient accélérées.
Selon cette logique, une baisse, même relative, de la saturation du sang en oxygène pourrait participer à une diminution de la vitesse d'utilisation de l'oxygène par le muscle.
Ce modèle est bien entendu général. Il doit être complété d'autres interactions que la seule quantité de substrats. Nous pensons notamment à l'acidité, au taux de dégradation de l'ATP, au rejet de CO2. qui influent sur la capacité de l'organisme à apporter l'oxygène aux muscles. [Ce dioxyde de carbone (CO2) que nous délaissons et qui pourtant pourrait être un élément majeur de la limitation de la consommation d'oxygène (hypothèse du biologiste français Henri Laborit)].
Quoi qu'il en soit, le schéma que nous déployons, à propos de l'oxygène, n'explique pas tout. Il dit simplement que toutes autres conditions étant égales, un apport plus important d'oxygène participe à une utilisation plus rapide.
Selon ce modèle, il ne conviendrait pas de chercher une quelconque limite unique et bien circonscrite à la capacité de l'organisme à utiliser l'oxygène.
A l'image de la capacité à produire un exercice physique intense, cette capacité serait le fait :
de données génétiques donnant un canevas susceptible de varier,
d'adaptation liées à la répétition des sollicitations (entraînement),
de lois physiques décrivant les phénomènes mis en jeu (exemple de la diffusion).
Ces trois données (gène, action de l'environnement et lois physiques) spécifieraient à un moment donné les vitesses maximales pouvant être atteintes par chaque transfert, par chaque réaction chimique. Sur ces données fixées à un moment donné, viendrait se greffer un équilibre dynamique qui serait le fait de :
différences de vitesses existant entre des réactions qui se suivent. Ces différences peuvent provoquer des accumulations de substrats qui correspondent à une limitation locale due aux systèmes chargés de leurs transformations.
de variations de vitesses allant dans le sens (1) d'une augmentation de la rapidité des réactions (ou des transferts) participants à réduire le surplus d'un élément et (2) de la baisse de vitesse de celles concourant à augmenter ce surplus. Au total, il s'agirait d'un ajustement des vitesses visant à réduire les différences de concentrations, les écarts à l'équilibre. Cet ajustement serait le fait :
de rétroactions positives ou négatives (voir physiologie/énergie) qui se joueraient entre les différents acteurs des réactions chimiques (ATP, transporteurs, CO2, acidité.).
Un tel modèle rend compte d'un fonctionnement général de l'organisme. Il explique aussi bien l'utilisation d'oxygène que la capacité de performance physique.
Si nous le suivons, les différents éléments qui composent le système de transformations énergétiques et d'échanges gazeux conditionnent tous plus ou moins la grandeur de VO2max et de l'effort pouvant être accompli. Bien entendu, ce modèle est moins facile à imaginer, moins aisé à contrôler et à modifier qu'une limitation bien localisée. Cependant, nous sacrifions à la simplicité en pensant nous approcher de la réalité du fonctionnement de notre organisme.
Toutefois, à des fins de simplifications, nous pouvons estimer que des limitations principales à l'utilisation d'oxygène peuvent être avancées. Ces limitations varient selon le niveau atteint par les personnes. Dans l'état actuel de notre réflexion, nous pensons pouvoir avancer - avec prudence - que les facteurs déterminants sont :
les possibilités du muscle pour les sujets sédentaires. Ces sujets ne présentent pas de limitation respiratoire ; leurs possibilités mécaniques maximales ne sont pas atteintes et ils n'ont pas de désaturation oxyhémoglobinée à l'exercice maximal. Enfin une altération de leur capacité de transport de l'O2 par un béta bloquant n'a pas d'effet sur leur VO2max.
le débit cardiaque pour les sujets dotés d'une bonne aptitude aérobie (VO2max entre 50 et 60ml/min/kg). Ces sportifs présentent une baisse de VO2max quand leur capacité de transport de l'oxygène est diminuée par un béta bloquant.
la respiration chez certains athlètes bien entraînés (VO2max supérieure à 60ml). L'apparition d'une désaturation du sang en oxygène à l'approche de l'exercice maximal, laquelle disparaît en cas d'apport d'oxygène supplémentaire, permet de penser à cette limitation respiratoire. Chez les athlètes très entraînés ne présentant pas de baisse du taux d'oxygène dans le sang, le débit cardiaque pourrait être le principal facteur limitant la consommation de ce gaz.
Les modèles "complexe" et "académique" ayant été abordés, nous pouvons terminer ce chapitre en parlant des effets de l'entraînement. Nous aborderons dans un premier temps l'ensemble des effets physiologiques aérobies produits par l'entraînement puis nous relaterons quelques expérimentations - certaines connues d'autres moins - ayant étudié les effets de l'entraînement sur les qualités aérobies des sportifs.