4 les filières énergétiques
 Présentation
Présentation
Dans le chapitre consacré aux qualités physiques de l'athlète , nous parlons de trois filières permettant de transformer l'énergie :
 la filière aérobie utilisant l'oxygène de l'atmosphère
la filière aérobie utilisant l'oxygène de l'atmosphère
la filière anaérobie acide produisant de l'acidité dans l'organisme
la filière anaérobie anacide n'utilisant pas d'oxygène et ne produisant pas d'acidité
Dans les pages précédentes (énergie 1 et 2), nous parlons de voies de transformation des glucides, lipides et protides. Or, dans les deux cas, nous disons que l'objectif est de fournir de l'énergie pour le mouvement ! Il y a vraiment de quoi se perdre dans ces méandres d'explications.
En fait, l'ensemble est plus simple qu'il n'y parait. Dans les deux cas, nous parlons de la même chose mais selon une approche différente. Les catégories servant de base à la réflexion ne sont pas les mêmes mais les processus qu'elles décrivent sont strictement identiques. Nous avons agit un peu comme si nous décrivions une maison en nous basant une fois sur les couleurs (le toit est brun, les murs sont beiges .) et une autre fois sur les formes (les pentes du toit composent un angle aigu, les murs constituent un cube.). Dans les deux cas, nous parlons de la même maison ; mais nous en parlons différemment. Alors que la description qui précède se base sur les substrats alimentaires, celle que nous entreprenons s'appuie sur la présence ou l'absence d'oxygène et d'acidité.
Commençons par la catégorie qui justement ne comprend aucun des deux éléments. Commençons par la filière dite "sans oxygène et sans acidité".
4.1 le système de mise en réserve d'énergie : une boîte à remplir
Peut-être êtes vous surpris de constater que le titre de cette partie n'est pas : la filière sans oxygène et sans acidité (ou sans lactate) ? Si nous avons choisi de ne pas reprendre ce vocable communément admis, c'est qu'il est bien mal adapté à ce que nous allons décrire.
Il est vrai que le système dont il va être question fourni de l'énergie aux muscles sans utiliser d'oxygène. Mais à part ça, le reste de l'appellation est totalement erroné.
1le système ne constitue aucunement une voie énergétique puisqu'il se limite à trois malheureuses réactions qui ne "s'emboîtent" pas complètement.
2sur les trois réactions, une produit de l'acidité
3les réactions ne prennent pas d'énergie aux substrats mais ne font que récupérer, mettre en réserve et donner aux muscles de l'énergie apportée par les deux seules voies énergétiques : la filière aérobie et la filière anaérobie.
Par commodité de langage, nous avons maintenu l'appellation de filière énergétique dans le secteur consacré à l'entraînement des qualités physiques .
Ceci étant dit, passons au premier des composés de mise en réserve de phosphates à haute énergie : l'ATP.
4.1.1 L'ATP (adénosine tri phosphate)
Nous l'avons dit à plusieurs reprises ; pour que le mouvement soit possible, l'énergie doit passer de sa forme alimentaire à une forme utilisable par le muscle. Or - et c'est là encore une tendance générale au vivant - l'énergie directement utilisable par un organisme est presque systématiquement portée par des composés phosphatés proches de celui utilisé par nos muscles. Ce composé porte le doux nom d'ATP (adénosine tri phosphate).
Pour courir, nous utilisons donc de l'ATP. Nous le coupons et récupérons l'énergie que cette scission libère. Le problème est que tout l'ATP du corps tient dans notre main et suffit à peine à un exercice d'une seconde.
Pour continuer à vivre au-delà de ce court répit, notre organisme doit se débrouiller pour reformer la molécule détruite. Toutes les réactions de dégradation de l'énergie des aliments sont orientées vers ce but. Elles concourent toutes à la réalisation de cette tâche.
Une somme considérable d'enzymes, de métabolites. participent à la kyrielle des réactions chimiques aboutissant à la restauration de la molécule d'ATP. Et elles y réussissent plutôt bien puisque à chaque instant ce sont des milliards de molécules d'ATP qui sont reconstituées. Si nous remplaçons une molécule d'ATP par un grain de sable. Ce travail chimique s'apparente à l'activité de milliers d'engins mécaniques qui, à chaque seconde, recouvriraient d'une couche de plusieurs mètres des centaines de stades de football.
Ces réactions, d'une multiplicité qui dépasse notre entendement, suivent pourtant des principes relativement simples. Nous évoquons ces règles de fonctionnement dans la page, nous les approfondirons par la suite. Elles s'appliquent aux processus qui suivent.
Dégradation et reconstitution de l'ATP
C'est en 1929 que Lohmann a découvert la molécule d'ATP. Cette molécule est constituée d'un composé complexe - l'adénosine - et de trois parties plus simples ; les groupements phosphate. Or, ce qui caractérise les liens entre deux phosphates, c'est l'énergie qui leur est associée. Pour établir une telle liaison une grande quantité d'énergie doit être apportée à la réaction. A l'inverse, lorsqu'elles sont cassées, les liaisons rendent toute leur énergie.
Au total, lorsqu'une liaison phosphate est rompue :
- un groupe phosphate est enlevé de la molécule d'ATP
- de l'énergie est produite (de 7 à 12 kcal selon Hultman et Karlsson)
- de l'adénosine diphosphate (ADP) et un phosphate inorganique (Pi) sont formés.
La réaction chimique associée est : ATP  ADP + P + énergie
ADP + P + énergie
ADP : adenosine diphosphate, P : phosphate
Comme nous le voyons dans le chapitre consacré au mouvement (physiologie/le mouvement), la réaction de dégradation de l'ATP est rendue possible grâce à des enzymes appelées ATPases. Différentes formes d'ATPases se retrouvent aux endroits de l'organisme où l'utilisation d'énergie est nécessaire : dans les parois cellulaires (ATPase Na+/K+), au niveau du réticulum (ATPase Ca2+) ; au niveau des têtes des filaments de myosine (ATPase myosine). Cette pluralité se retrouve encore à l'étage inférieur puisque les enzymes de la contraction musculaire (ATPase myosine) présentent plusieurs configurations. En fonction du type d'enzyme intervenant dans la dégradation de l'ATP, les caractéristiques de la réaction - comme sa vitesse par exemple - sont modifiées. Nous avons souligné les possibilités d'adaptation associées à une telle diversité (secteur tout sur l'entraînement).
Remarque
La réaction que nous avons donnée est simplifiée pour trois raisons.
1Dans la cellule, la molécule d'ATP est présente principalement sous la forme d'un complexe magnésium : MgATP2-.
2La réaction de dégradation se fait par l'intermédiaire de l'eau et prend le nom d' hydrolyse.
3En plus de l'énergie, elle produit de l'acidité sous la forme d'un proton H+. Une fois ces approximations rétablies, la réaction devient : MgATP2- + H2O HPO42- + MgADP- + H+ + énergie
HPO42- = Pi : phosphate inorganique
Cette petite remarque ne pourrait être qu'une précision. Mais, elle établit ce fait important : la dégradation de l'ATP produit de l'acidité.
La réaction de dégradation de l'ATP est l'étape ultime précédent la contraction musculaire. Ce qui se passe avant cette réaction est de l'ordre de la chimie, ce qui la suit est, avant tout, de l'ordre de la mécanique (voir physiologie/.
Deux autres réactions basées, elles aussi, sur la mise en réserve de l'énergie par des composés phosphates viennent seconder l'ATP.
4.1.2 reconstituer l'ATP grâce aux composés phosphate
La première de ces réactions s'appuie sur la créatine phosphate (CP) ou phosphocréatine (PC).
CP (ou PC)
Réaction : PC + ADP ATP + C
C : créatine
La réaction est catalysée par l'enzyme CPK (créatine-phospho-kinase)
Si la phosphocréatine peut régénérer un ATP c'est que l'énergie portée par sa liaison phosphate est au moins aussi grande que celle portée par l'ATP (Cf page précédente .
La créatine joue le rôle de réservoir supplémentaire à énergie. Son action est complètement inféodée à l'ATP. Elle est là pour apporter l'énergie immédiate permettant sa synthèse. Tout juste dégradé, l'ATP est rétabli à partir de l'ADP et du phosphate (P) grâce à l'énergie libérée par la dégradation des réserves de CP.
La phosphocréatine se reconstitue au repos, lorsque l'ATP est redevenu abondant, selon la réaction précédente s'effectuant en sens inverse : ATP + C CP + ADP
L'énergie nécessaire à la synthèse de CP est donc fournie par la dégradation de l'ATP. Ce processus a lieu lors de la récupération suivant l'exercice. A ce moment, l'énergie nécessaire à la reconstitution de l'ATP provient directement de la dégradation des aliments par les deux voies énergétiques.
Enfin, la dernière réaction du système phosphate passe par la création d'un adénosine mono phosphate
AMP
Réaction : 2 ADP ATP + AMP
1,2 et 3 phosphates ! Nous avons fait le tour simplifié du système.
Cette troisième réaction a exactement le même statut que celle basée sur la créatine phosphate. Au niveau énergétique, elle est certainement plus marginale que cette dernière. En revanche, elle permet d'augmenter le rapport ATP/ADP qui - nous le verrons - agit comme régulateur des transformations énergétiques de l'organisme ( la régulation).
Après avoir décrit le système phosphate, voyons dans quelles mesures il peut être modifié par l'entraînement.
4.1.3 le système phosphate et l'entraînement
Parlons, dans un premier temps, de l'évolution des taux des éléments phosphate au cours de l'exercice physique.
L'exercice
Durant les premières secondes d'un exercice musculaire intense, l'ATP se maintient à un niveau relativement constant tandis que la phosphocréatine (PC) diminue régulièrement. S'il en est ainsi, c'est qu'elle est utilisée pour régénérer l'ATP. Moins d'un tiers de seconde après le début de l'exercice, la PC est déjà à l'ouvre pour régénérer l'ATP. Cette dégringolade du taux de PC que cette activité de soutien provoque est rapidement suivie de celle du taux d'ATP. Après quelques secondes de sprint, le taux d'ATP atteint sont niveau minimal à savoir 80% de sa valeur de repos. (Il faut savoir qu'une baisse de moitié des réserves d'ATP est sans doute suffisante pour causer la mort de la cellule.) Les taux d'ATP et surtout de PC sont alors bien déprimés. Cet état s'accompagne d'une fatigue ressentie par les athlètes (Hultman, 1991 pour la PC). Pourquoi alors ne pas tenter de palier à cette fatigue, et à la perte d'efficacité qui va avec, en augmentant le niveau de créatine dans le muscle ?
Créatine et exercice
La créatine est synthétisée par le corps -à partir des acides aminés arginine, glycine et méthionine - et apportée par l'alimentation carnée (viande).
L'idée d'approvisionner l'organisme en créatine pure dans l'espoir d'augmenter le niveau dans les muscles fut testée. Cette idée mise en pratique dans de nombreux sports fait, aujourd'hui, l'objet de vifs débats.
Si nous nous limitons au domaine de la performance, les études ont montré que l'ingestion répétée de créatine par petites doses augmente sa concentration jusqu'aux limites supérieures habituellement observées chez l'homme. Parallèlement, la force musculaire des sujets est augmentée dans des proportions de l'ordre de 5 à 10%.
En dépit de cette amélioration, les participants de cette étude peuvent toujours s'accrocher pour réussir à rattraper le champion de la créatine qu'est le homard. Ce dernier possède un métabolite phosphate, quasiment identique à la PC, en quantité quatre fois supérieure. Cette disposition lui permet d'effectuer des sprints qui feraient pâlir de jalousie un sprinter averti. Si toutefois, les sujets de l'étude s'entichaient à continuer leur ingestion de créatine à des fins de performance, ils devront prendre garde à leur santé. Une étude vient d'établir que le supplémentation en créatine augmenterait les risques de cancer.
Mais revenons aux procédés plus "propres".
L'entraînement
L'entraînement de vitesse courte ne permet pas d'augmenter les taux d'ATP et PC. Il permettrait uniquement d'accroître la déplétion en PC. En d'autres termes, l'entraînement a pour effet de vider un peu plus le réservoir PC. En plus de cet effet physiologique, il augmente la force musculaire.
Par ailleurs, l'entraînement à la vitesse prolongée (courses de 30 secondes) augmenterait également l'activité des enzymes activant la dégradation du PC et de l'ATP. Il aurait donc pour effet d'augmenter la vitesse du flux énergétique (Costill et Al, 1979).
Nous en avons terminé du "système phosphate" de mise en réserve de l'énergie. Nous aurons compris, que les phosphates peuvent être comparés à une petite colonie chargée de récupérer dans une boîte commune l'énergie en provenance de la dégradation des substrats puis de la distribuer aux muscles en cas de besoin. Le chef de la colonie, le seul qui puisse parler au muscle, c'est bien sûr l'ATP. C'est également par lui, que les deux filières énergétiques vont pouvoir apporter leur énergie aux muscles.
4.2 la filière aérobie
Si nous nous rapportons au schéma figurant les réactions énergétiques , nous constatons que nous avons séparé d'une barre la partie haute des transformations de la partie basse.
Cette ligne représente la membrane de ce que nous avons appelé la centrale énergétique de la cellule, à savoir, la mitochondrie.
La mitochondrie
Les mitochondries ont une longue histoire. A l'origine, elles seraient des bactéries qui auraient colonisé d'autres organismes vivants. Vivant en symbiose avec leur hôte, elles se seraient spécialisées dans la transformation d'énergie d'origine aérobie.
Depuis des millions d'années, elles ont conservé une partie de leur patrimoine génétique. Actuellement encore, alors que toutes les cellules de notre corps possèdent la même information génétique, les mitochondries restent à part. Elles ont leur propre matériel génétique, lequel n'est apporté que par la mère. Nous devons donc notre patrimoine génétique aérobie à notre mère uniquement.
Après le contenant, revenons au contenu.
La partie inférieure du schéma présente de manière extrêmement simplifiée les réactions se déroulant au sein de la mitochondrie. Ce sont elles qui apportent l'immense majorité de l'énergie dont notre organisme a besoin.
Si vous souhaitez connaître plus spécifiquement les transformations aérobies - c'est à dire utilisant l'oxygène comme accepteur final d'électrons et de protons - vous pouvez vous rendre au chapitre aérobie (physiologie/aérobie). Nous y traitons de tout ce qui se rapporte à l'oxygène : son accumulation dans l'atmosphère il y a plus de 2 milliards d'années, son parcours le menant de l'air ambiant à l'intérieur de nos cellules en passant par les poumons puis le sang, son utilité à l'intérieur de chacune de nos cellules, son utilisation par le corps au cours de la course...
Dans la partie qui suit, nous nous attachons à présenter la partie haute du schéma. Cet ensemble de réactions qui précède les transformations aérobies et que nous nommons anaérobie acide.
4.3 la filière anaérobie acide
Nous appelons filière anaérobie acide la suite de 10 réactions assurant la transformation du glucose sans intervention de l'oxygène. Ces réactions constituent la partie haute du schéma présenté précédemment.
Pour le sportif, cette filière apporte un certain nombre d'avantages mais aussi d'inconvénients. Commençons par un sérieux atout, sa rapidité.
4.3.1 Les "plus" de la filière anaérobie acide
Premier avantage de la filière acide, sa rapidité de mise en action. Au niveau temporel, un substrat passe d'abord les étapes anaérobies puis pénètre dans la "sphère à oxygène". Cet agencement a deux conséquences. La première est que les deux filières sont imbriquées. L'anaérobie prépare et nourrit l'aérobie. La seconde - qui nous intéresse plus directement dans ce paragraphe - est que cette prime position rend l'intervention de la filière anaérobie extrêmement rapide.
Les premières études sur la physiologie de l'effort avaient conclues que les trois filières se succédaient dans le temps. Selon ce modèle dit de Howald, proposé il y a une cinquantaine d'années, la filière appelée anaérobie alactique (système phosphate) s'activait en premier. Après 20 secondes d'exercice, la filière anaérobie acide (anaérobie lactique) s'activait à son tour pour palier l'épuisement des réserves d'ATP et de PC. La filière acide était à son tour relayée après deux minutes d'exercice par les transformations aérobies.
Nous savons maintenant que dès la première seconde de contraction d'une fibre musculaire, les transformations d'origine anaérobie acide participe de manière aussi marquée au renouvellement de l'énergie que les composés phosphates (anaérobie anacide) (Hultman et Sjöholm, 1983).
Cette rapidité s'explique, en partie, par le fait que cette filière n'a pas besoin d'oxygène pour fonctionner. Cette caractéristique constitue le deuxième atout de cette voie énergétique. Elle n'est pas dépendante de l'oxygène que l'organisme tire de l'atmosphère.
Enfin, la filière anaérobie dispose d'une vitesse de transformation des substrats assez élevée (cf tableau présenté plus bas : vitesse de la glycolyse. C'est grâce à cette vitesse de transformation que nous pouvons courir à des allures supérieures à la vitesse maximale aérobie.
Voyons, à présent, le revers de la médaille.
4.3.2 Les "moins" de la filière acide
Le premier problème de la filière anaérobie acide est qu'elle ne peut utiliser que le glycogène ou le glucose pour fournir de l'énergie. Or le rendement de la transformation anaérobie du glucose est extrêmement faible. La suite de 10 réactions qui compose la voie acide produit un total net de seulement deux ATP par molécule de glucose. Par comparaison, la seule partie aérobie de dégradation du glucose fournit 34 ATP.
La conséquence est double :
1 la quantité totale d'énergie pouvant être délivrée est assez faible.
2Une dépendance trop grande au métabolisme anaérobie causerait rapidement un épuisement des réserves en glucose et en glycogène. Par exemple, dans une course de 10 000m, la conversion du glycogène en lactate fournie moins de 5% de l'ATP nécessaire à la course tout en faisant chuter d'un tiers les réserves en glycogène.
Le deuxième problème est l'accumulation des protons H+. La glycolyse anaérobie extrait du glucose les protons H+ accompagnés de leur électron. Quand la demande énergétique est limitée, la totalité de ces protons est prise en charge par des transporteurs (T-H+ - en orange sur le schéma page précédente) qui les conduisent au niveau de la chaîne respiratoire (physiologie. En revanche, lorsque les besoins en énergie sont élevés, la glycolyse accélère son rythme ; les transporteurs tentent d'en faire autant mais sans parvenir à la même efficacité. Ils n'ont plus le temps de faire les allers et retours entre la glycolyse et la chaîne respiratoire. Les protons qu'ils devraient prendre en charge s'accumulent dans la cellule. Et qu'y font-ils ? Ils entraînent l'acidité du milieu. Les responsables de l'acidité de la cellule sont les protons H+.
Acidité, protons H+ et lactate
Les responsables de l'acidité de la cellule sont les protons H+ et pas, comme il est souvent dit, l'acide lactique. Ce composé ne fait que palier à la différence de vitesse qui existe entre la filière anaérobie et la transformation aérobie de l'énergie.
Pour comprendre ce qui se passe, nous devons entreprendre un petit détour. Le dernier métabolite de la voie anaérobie est le pyruvate. Habituellement, ce composé pénètre dans la mitochondrie où il est dégradé au cours du cycle de Krebs. Mais, là aussi, le cycle de Krebs avance moins vite que la glycolyse anaérobie ne peut le faire. Si bien que lorsque la filière anaérobie tourne à haut régime, le cycle aérobie ne peut suivre, le pyruvate s'accumule.
Or, quand il s'accumule, le pyruvate capte des protons H+ présents dans la cellule. Ce faisant, il devient de l'acide lactique. Or cet acide a tendance à se dissocier en ion lactate et proton H+. En d'autres termes, il redonne son proton.
Voilà sa seule responsabilité. Il prend et redonne l'acidité. Finalement, nous lui prêtons tous les maux du sportif et pourtant il n'en est que l'expression (à ce propos voir.
NB : A propos de proton H+, signalons à titre de remarque complémentaire, que nous employons le terme H+ pour faciliter le discours. En fait, le proton hydrogène H+ n'existe pas à l'état libre. Il se combine à l'eau pour donner l'ion hydronium (H3O+). Cette association entre l'eau et l'ion hydrogène ne change rien à ce que nous avons dit de lui.
Si vous êtes intéressé par l'acide lactique, par son parcours dans l'organisme, par son évolution pendant l'exercice, par son rapport avec la performance en course à pied. vous pouvez vous rendre au chapitre que nous lui consacrons.
Mais revenons à notre acidité. Dans l'organisme, l'acidité produit des effets qui ne sont pas toujours avantageux pour le sportif.
elle limite l'activité de la glycolyse anaérobie.
elle bloque la contraction musculaire et limite l'utilisation de l'ATP
elle favorise la détérioration des tissus
elle engendre une douleur difficile à soutenir
Bien que négatives pour la performance, ces adaptations permettent, pour l'essentiel, de préserver l'organisme. Si la glycolyse continuait, l'acidité augmenterait rapidement jusqu'à un point où toutes les enzymes s'arrêteraient de travailler et où toutes les cellules mourraient. La douleur, la réduction de la vitesse des réactions et de la contraction préviennent une telle issue. Elles agissent comme une sécurité permettant de protéger l'organisme. Un des moyens utilisé consiste donc à contrôler la vitesse de la filière.
4.3.3 Vitesses des filières
Le tableau qui suit donne la vitesse de dégradation des substrats énergétiques.
|
Réaction
|
Vitesse (mol/min/kg de muscle) |
|
Dégradation de l'ATP
Dégradation du PC
Glycolyse anaérobie
Dégradation aérobie du glucose
Dégradation aérobie des lipides |
156
120
73
38
20
|
Vitesses de dégradation des substrats (tableau ci-dessus et graphique ci-dessous). Le schéma est reproduit dans le chapitre consacré à l'acide lactique (physiologie). Les couleurs du schéma font référence aux zones d'allures de course
|
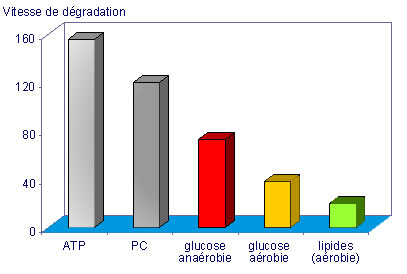
Nous pouvons constater que la glycolyse anaérobie dispose d'une vitesse de transfert de l'énergie intermédiaire entre celle du système phosphate (ATP-PC) et celle de la voie aérobie.
Il semble que le "maillon faible" de cette voie, celui qui détermine la vitesse de l'ensemble, se trouve au niveau de la troisième réaction. Cette réaction transforme le fructose 6-phosphate en fructose 1,6-diphosphate. Elle est sous le contrôle d'une enzyme appelée phospho-fructo-kinase (PFK). C'est peut-être le taux de cette enzyme qui limiterait la vitesse de toute la filière anaérobie.
Si nous faisons le lien avec l'entraînement, ce que nous avons appelé la puissance acide serait limité par cette PFK. Nous verrons qu'un des effets de l'entraînement est justement d'accroître la quantité et l'activité de cette enzyme.
En dépit de l'importance que cette information peut avoir pour le coureur de 400m, elle n'est pas majeure pour le spécialiste de demi-fond et de fond. La capacité de ce dernier à courir vite est beaucoup plus déterminée par les différences de vitesses entre les filières énergétiques.
Différences de vitesses
Que ce soit au niveau du rapport entre les transporteurs et les protons H+ ; de la filière anaérobie et aérobie. les différences de vitesses se retrouvent à tous les niveaux des transformations énergétiques. Dans le tableau précédant, nous pouvons constater, par exemple, que la glycolyse sans oxygène peut aller près de deux fois plus vite que la dégradation du glucose avec oxygène. La conséquence de la différence de vitesse entre deux systèmes est l'accumulation sur place du métabolite faisant le lien entre eux.
Le déséquilibre qui se crée entre les métabolites, suite à l'inégalité des vitesses, est le point clé de la régulation des transformations énergétiques ; régulations dont nous parlerons juste après avoir vu les effets que produit l'entraînement sur la filière anaérobie acide.
4.3.4 filière acide et entraînement
Dans le secteur consacré à l'entraînement, nous différencions l'entraînement de type puissance acide de celui de type capacité. L'entraînement de type puissance est associé à des distances courtes, proches de 300m, réalisée à vitesse maximale alors que le versant capacité sollicite la filière plus longtemps. Les effets physiologiques de ces deux modalités d'efforts seraient différents.
La puissance acide
Elle augmenterait la quantité totale d'énergie libérée en absence d'oxygène. L'objectif est d'ouvrir plus grand le robinet acide. Ce type de travail provoque d'importants taux d'acide lactique et une acidité record. Il interviendrait en priorité sur l'aptitude du muscle à produire de l'énergie malgré l'acidification grandissante. Suite à un entraînement de la filière acide, la quantité d'enzymes (phosphorylase, PFK, LDH...) assurant les processus de dégradation des substrats augmenterait de manière marquée (Sharp et Al, 1986).
Un tel accroissement ne se retrouverait pas suite à un entraînement plus court. Des répétitions de 5" permettrait cependant d'augmenter l'activité des enzymes de la glycolyse anaérobie.
La capacité acide
Le travail de capacité acide permettrait d'améliorer la diffusion de l'acidité (expulsion en dehors de la cellule) et sa métabolisation (neutralisation des protons H+). Il s'opposerait donc à la montée acide. En conséquence, le niveau acide atteint et le taux d'acide lactique accumulé dans le muscle seraient moins élevés pour un même niveau de production. Cet effet est connu sous le terme de capacité tampon. Huit semaines d'entraînement à base de répétitions d'exercices de 30 secondes augmenterait de 12 à 50% le système tampon de l'acidité (Sharp et al, 1986).
Ce chapitre sur l'entraînement étant terminé, passons à la régulation des transformations énergétique au sein de la cellule. Elles vont nous permettre de comprendre pourquoi - entraînement ou pas - la vitesse de notre course sera toujours limitée.